
Processus moléculaire de l'expression génétique
Molécule d'ADN
Une molécule d'ADN a deux brins polynucléotidiques qui forment une double hélice antiparallèle. L'unité monomère d'un brin d'ADN est un nucléotide qui consiste en une base, un sucre de désoxyribose et un groupe phosphate. Les nucléotides successifs d'un brin d'ADN sont liés par des liaisons phosphodiester et les deux brins d'ADN sont maintenus ensemble par des liaisons hydrogène entre des paires de bases complémentaires spécifiques. Au cours de la réplication, qui est médiée par un certain nombre de protéines différentes, y compris des ADN polymérases, chaque brin d’ADN joue le rôle de matrice pour la production d’un brin complémentaire.
Protéines
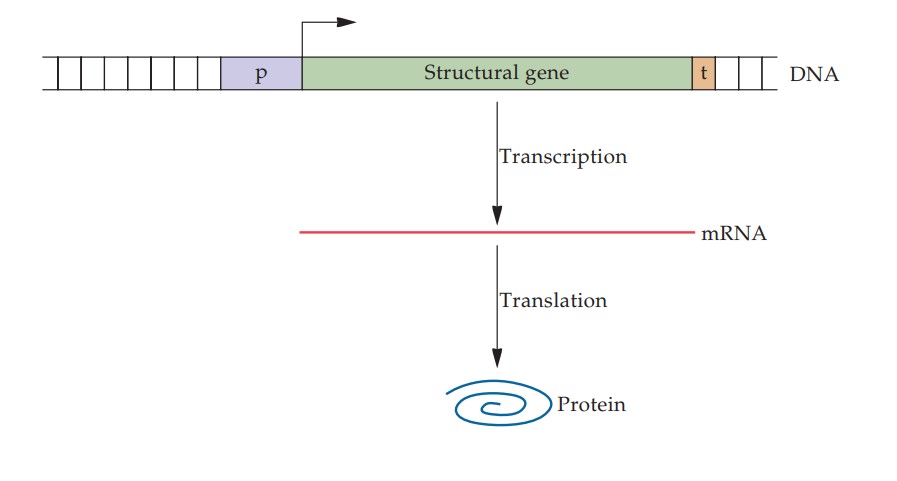
Les protéines sont essentielles au maintien de toutes les fonctions biologiques. Une protéine consiste en une séquence spécifique d'acides aminés liés par des liaisons peptidiques. La séquence d'acides aminés d'une protéine est codée dans l'ADN. Le processus de décodage de l'information génétique est effectué par des molécules d'ARN, y compris l'ARNm, l'ARNt et l'ARNr; diverses enzymes; et un assortiment de facteurs protéiques. Tout l'ARN est transcrit à partir de l'ADN.
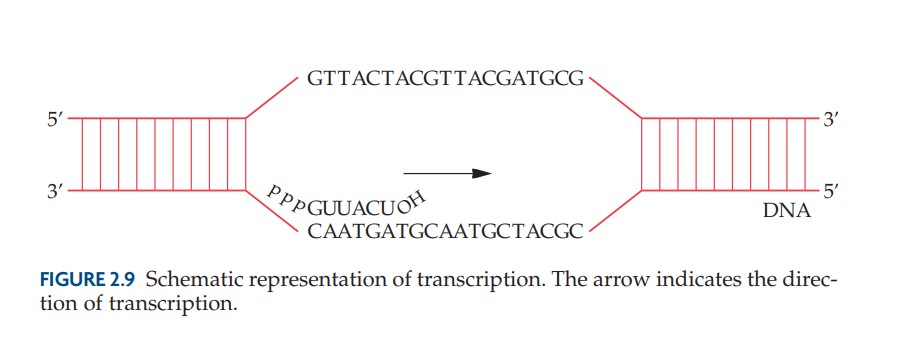
Transcription
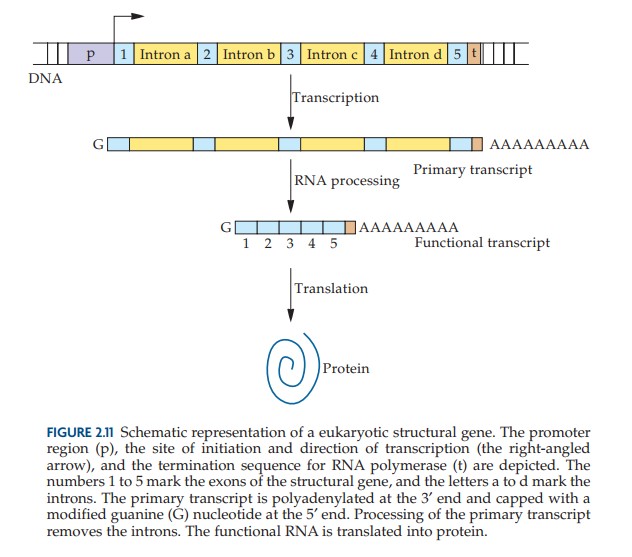
Des séquences d'ADN combinées à des facteurs protéiques garantissent que la transcription est initiée à un point de départ précis, que le brin approprié est utilisé comme matrice et que la terminaison se produit sur un site nucléotidique spécifié. Dans les organismes eucaryotes, la plupart des gènes structurels consistent en des régions codantes (exons) séparées par des segments non codants (introns). Les transcriptions primaires contiennent à la fois des exons et des introns. Cependant, un système de traitement supprime les introns et associe les exons, dans le bon ordre, pour former un ARNm fonctionnel. Un ARNm porte le code pour la séquence d'acides aminés d'une protéine.
Traduction
La traduction de l'ARNm pour produire une protéine se produit sur des ribosomes composés d'une grande et d'une petite sous-unité, chacune contenant un ARNr et un grand nombre de protéines spécifiques. La traduction chez les procaryotes est initiée par la réunion d'un ARNm avec une petite sous-unité ribosomale. Suite à l’appariement des bases complémentaires codon-anticodon, l’ARNt initiateur, fMet-tRNAfMet, se fixe au complexe ARNm – petit sous-unité ribosomique, qui se combine ensuite à la grande sous-unité ribosomale pour former un complexe d’initiation. La traduction chez les eucaryotes est initiée par la combinaison d'un ARNt initiateur unique qui porte l'acide aminé méthionine Met-tRNAMet avec une petite sous-unité ribosomique, puis par l'enfilage d'un ARNm à travers le complexe initiateur d'ARNt – petit ribosome jusqu'à la première AUG. séquence dans les paires d'ARNm avec l'anticodon de l'ARNt initiateur. La grande sous-unité ribosomale rejoint le complexe ARNm initiateur-petit ribosomal pour former un complexe d'initiation prêt à la traduction de la séquence de l'ARNm. Après la formation du complexe d'initiation, la phase d'élongation de la traduction est très similaire chez les procaryotes et les eucaryotes. Les trois nucléotides suivants dans la paire d'ARNm avec l'anticodon d'un ARNt qui porte son acide aminé spécifique. Le premier acide aminé, la méthionine, est clivé de l'ARNt initiateur et relié au second acide aminé par une liaison peptidique. L'ARNt initiateur «vide» est éjecté du ribosome, le complexe ribosome se déplace et l'ARNt auquel le peptide en croissance est attaché occupe le site laissé vacant par l'ARNt initiateur éjecté. En conséquence du décalage (translocation), le codon suivant de l'ARNm est disponible pour se lier à l'anticodon approprié d'un ARNt qui porte son acide aminé spécifique qui sera joint au peptide en croissance. En répétant ces étapes, un polypeptide avec une séquence spécifique d'acides aminés est formé. La traduction est terminée lorsque l'un des trois codons d'arrêt est rencontré dans l'ARNm d'un ribosome. Un facteur de terminaison, plutôt qu'un ARNt, reconnaît le codon d'arrêt et la liaison entre le dernier ARNt et la chaîne d'acides aminés achevée est clivée, ce qui provoque la libération de l'ARNt, de l'ARNm et de la protéine complétée.
Régulation de la transcription
Seuls les ARN et les protéines indispensables au maintien des fonctions cellulaires de routine sont synthétisés en continu. Pour conserver les ressources cellulaires, la transcription des gènes restants ne se produit que lorsqu'une protéine particulière est requise et est désactivée lorsque la protéine n'est plus nécessaire. Chez les procaryotes, la transcription est initiée par la liaison de l'ARN polymérase aux éléments -10 et -35 de la région promotrice d'un opéron. Les protéines régulatrices qui se lient aux séquences d'opérateurs dans et autour de la région promotrice contrôlent l'activité de l'ARN polymérase au niveau du promoteur et contrôlent ainsi l'initiation de la transcription. Les répresseurs empêchent l'initiation de la transcription en bloquant la liaison de l'ARN polymérase au promoteur ou le mouvement le long de l'ADN, tandis que les activateurs améliorent la liaison de l'ARN polymérase à une séquence du promoteur. Les activités des protéines régulatrices sont contrôlées par de petites molécules effectrices qui augmentent ou diminuent leur liaison à la séquence de l'opérateur. Chez les eucaryotes, l'ARN polymérase II, qui transcrit les gènes structurels, se lie à un ensemble de protéines appelées facteurs de transcription qui se lient, en séquence, à une séquence TATA d'une région promotrice. D'autres facteurs de transcription qui se lient à des éléments d'ADN de gènes structurels eucaryotes sont responsables de l'activation ou de la désactivation de la transcription. L'expression des gènes eucaryotes est également influencée par la conformation locale de l'ADN chromosomique. Les régions fortement compactées par des protéines spécifiques associées à l'ADN ne sont généralement pas transcrites, alors que les régions plus lâchement emballées contiennent des gènes actifs sur le plan transcriptionnel.
Exportation des protéines
Les procaryotes et les eucaryotes disposent de systèmes spécialisés pour l'exportation de protéines à travers une membrane cytoplasmique. Les protéines procaryotes sécrétées ont une séquence d'acides aminés à leurs extrémités N-terminales qui cible la protéine soit vers la voie de sécrétion générale, soit vers des complexes protéiques plus spécialisés transportant des protéines spécifiques. Les protéines eucaryotes à sécréter sont synthétisées sur des ribosomes associés au réticulum endoplasmique et sont d'abord sécrétées dans la lumière du réticulum endoplasmique via une séquence signal N-terminale, où elles sont clivées, repliées et modifiées chimiquement. Après traitement ultérieur dans l'appareil de Golgi, les protéines sont transportées vers la membrane cytoplasmique dans des vésicules membranaires et, après la fusion de la vésicule et des membranes cytoplasmiques, sont libérées dans l'environnement externe.
{kind=link}
0 Commentaires